A new paper provides evidence that certain RNAs associate with centrosomes and may represent a centrosomal RNA genome. Furthermore this potential genome includes an enzyme that could copy the centrosomal associated RNA. But first some background.

For many years, there have been scientists interested in the mysterious entity called the centrosome. Often refered to as the black hole of the eukaryotic cell, these intracellular structures (
see red dots) are often located next to the nucleus (
blue), where the microtubule cytoskeleton (
green) converges (called the microtubule organizing center).
Another fact for you: the primary cilium, a long skinny cellular extension
found in most cells, grows off of one of the centrosomes. This primary cilium is probably involved in some sensing mechanism. In fact in some cells (such as your rods and cones) the primary cilium is where most of the cellular sensory apparatus is located. Mutations that result in short primary cilliums cause kidney problems and probably involve a missensing of some property of fuid within the kidney.
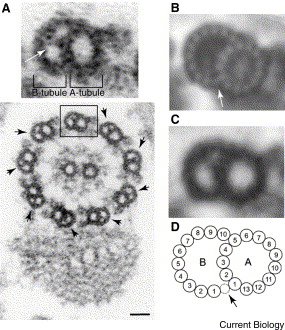
Centrosome are composed of two centrioles, barrel like structures made mostly of tubulin + other more unique proteins (see image below the fold). The number of centrosomes per cell is tightly regulated and the duplication of centrosomes is quite bizarre. You start off with a single centrosome. At some point (S phase?) the two centrioles come apart and new (or "daughter") centrosomes seem to grow off of the old (or "mother") centrosomes. Very reminiscent of
semi-conservative replication of DNA. But it get stranger.

If you watch the centrosomes throughout cell division, right before cytokinesis (the physical cleavage of two daughter cells) occurs, one of the two centrosomes from one of the two cells, migrates to the cleavage furrow and (in the words of
Michel Bornens) gives it a "kiss". This event is thought to stimulate the final braking off of the two daughter cells.
Then about 5-6 years ago
Alexey Khodjakov performed a remarkable experiment. He labeled the centrosomes with a green fluorescent protein (I think it was GFP-gamma-tubulin) then blasted them away with a laser. After centrosome duplication he blasted one of the two centrosomes and got cells with lopsided resulting mitotic spindles. The microtubules one side converged on a centrosome while the microtubules on the other side having no centrosome, became unorganized. Incredibly if he ablated both centrosome,
the cells could form a spindle (mostly due to the fact that motor dynein can focus microtubules without centrosomes). The cells then progressed through mitosis and even divided (although sometimes they were trapped in cytokinesis - see Bornens' kiss). After division they got a huge surprize,
the cells stoped dividing further. They discovered that centrosomes are needed to
initiate a new round of DNA synthesis (or the G1-S transition). So just before DNA is copied, a cell asks itself "is my centrosome okay?" If the answer is yes => proceed with DNA copying. If not => stop cell division. You can advance through this "check point" if you get rid of p53, a key protein that regulates the cell cycle.
But why all the fuss? What is so special about the centrosome? Why are cells checking it? Why is it kissing stuff?
Why? Michel Bornens came up with this
individuation theory. Centrosomes mark an individual cell. At certain points, cells check up on their centrosome to check on their individualistic identity. If something is wrong, such as improper cell division, cells can sense their non-individual state and activate programed death and/or other drastic measures. Screwed up cells (such as cancer cells) have all sorts of centrosome defects ... many have an overabundance of centrosomes and centrosome associated proteins suck as pericentrin.
 And there's more!
And there's more! Many years ago,
Lynn Margulis (famous for her theory of the
endosymbiotic origin of mitochondria) proposed that centrosomes originated from the fusion of a
spirochaete-like-organism (right, image of a spirochaete) and the prototypical eukaryotic cell. The spirochaete may have helped the host cell to move around (I never found this explanation very satisfying - the spirochaete must have offered some other benefit). In support of this idea, various publications in the 80s claimed that the centrosome contained its own nucleic acid, just like mitochondria and other products of symbiosis, but these experiments were never definitive.
Now out of
Bob Palazzo's lab (no relation ... see extra comment at the end) Mark Alliegro's Lab (
click here for a comment on this change) has come up with a truly remarkable finding. His group purified centrosomes from clam oocytes ... for some reason clams have huge centrosomes and thus makes this experiment a whole lot easier. Along with the centrosomes came ... certain specific RNAs. But wait, people have claimed this in the past, and one of the enriched RNA was 18s ribosomal RNA, perhaps it was contamination. On the other hand the enrichment was for only certain RNAs (five in this paper). Besides the 18s rRNA, the other four RNAs were not found in any genomic database. (
For you RNA enthusiasts these have now been called cnRNAs - centrosomal RNAs.)
And one of these cnRNAs is
QUITE intriguing. It encodes a protein that has similarity to 'RNA-dependent nucleotide polymerase'. Furthermore, this RNA probably gets translated into protein. (
Warning: technical jargon comming up) The group maid antibodies against a peptide derived from the hypothetical protein, and found an immunoreactive band in lysates from oocytes and adult clam (end jargon).
So let me rephrase this.
Clam centrosomes contain RNAs not found in any genome (so far), and one of these RNAs encodes an enzyme that could potentially copy RNA? Holly replication, Batman!So could it be? Centrosomes have their own genome (an RNA genome at that?) Wow!
Ref:
Mark C. Alliegro, Mary Anne Alliegro, and Robert E. Palazzo
Centrosome-associated RNA in surf clam oocytes
PNAS (2006) 103:9034-9038(Note: This whole Palazzo/centrosome thing is very quirky. He knows of me, I know of him, but we've never met. He has worked in the centrosome field for quite a while, I've published papers on a related structure, the microtubule organizing center. At meetings, some people think I have his long publication records, others who know Bob Palazzo well, ask me whether I'm his son -- he in fact does have a son named Alex. I almost went into the centrosome field by going to Steve Doxsey's lab (great guy!) but just couldn't live in Worcester. Now I'm studying (in part) RNA ... and Bob Palazzo is into cnRNA ... I know ... very strange.)